
 发布展会
发布展会
 发布资讯
发布资讯



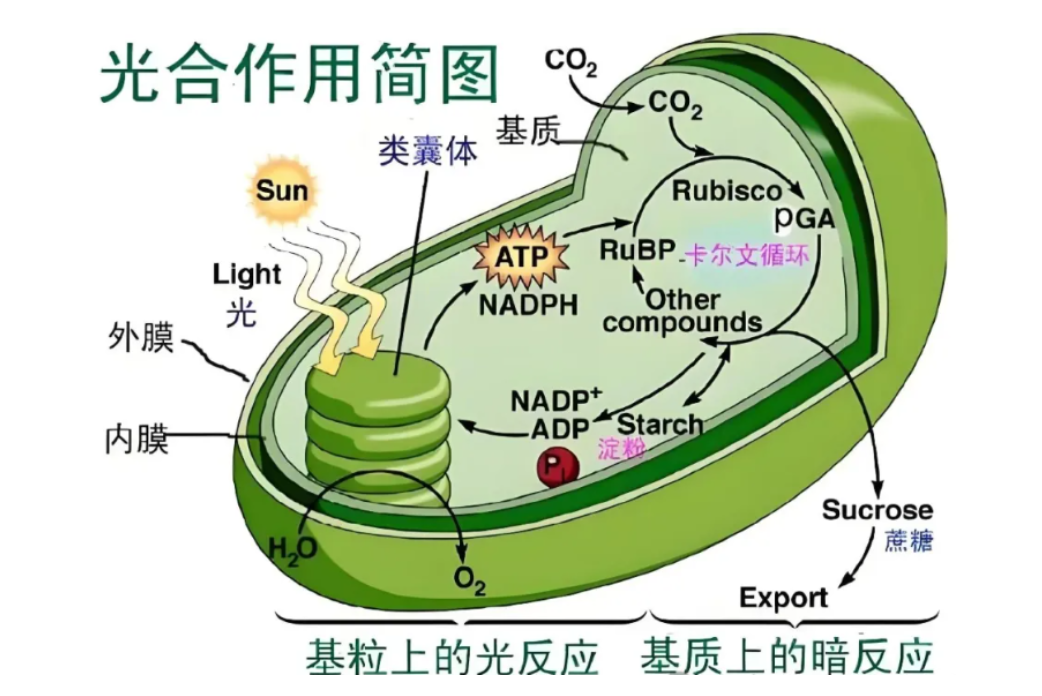
光合作用是自然界中最壮观的化学反应之一,它不仅支持着地球上大部分生命体的生存,还对维持地球的生态平衡起着至关重要的作用。本文将深入探讨光合作用的各个细节,从光合作用的基本结构到复杂的生化过程,再到光合作用在不同植物中的独特表现,力求全面展现这一神奇的生命现象。
光合作用的结构基础
光合作用的发生场所主要是植物细胞内的叶绿体。叶绿体中,类体膜是光合作用光反应的舞台,它富含蛋白质、脂类和色素,尤其是叶绿素和类胡萝卜素,这些色素负毒吸收太阳光能。
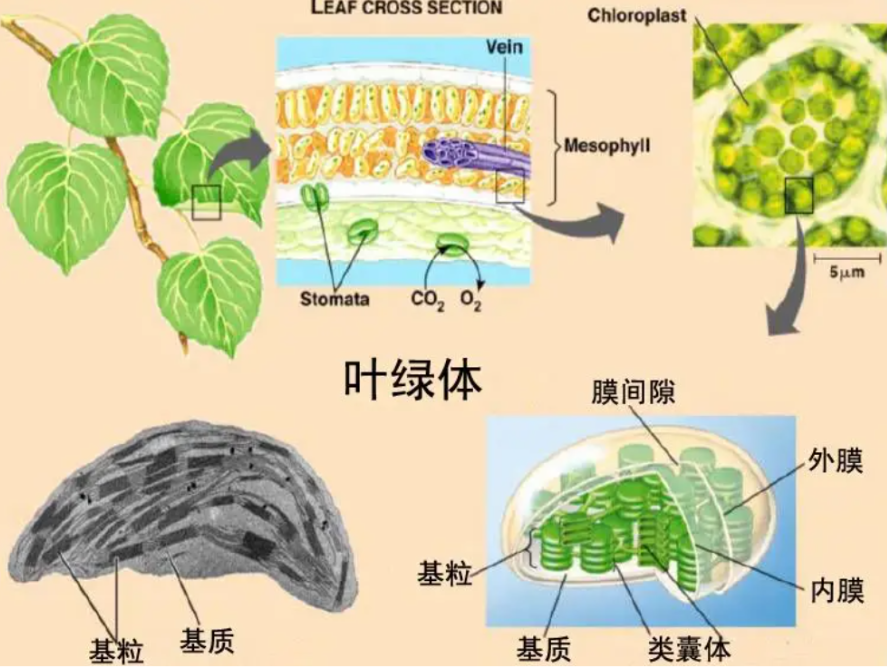
类囊体膜的流动性得益于其中含有较高的不饱和脂肪酸,这让光反应不易受温度变化的影响。光反应中心的色素分子,如叶绿素a,在类囊体膜上排列成复杂的蛋白复合体,其中包含200至300个天线色素分子,共同捕获光能。
二、光合作用的光反应
光反应发生在类囊体膜上,当叶绿素分子吸收光能后,从基态跃迁至第一单线态,随后释放出激发态电子,引发一系列电子传递事件

光系统I(PSI)和光系统|(PSI)是两个光反应中心,它们各自含有中心色素分子,PSll的中心色素为P680,而PSl的中心色素为P700。PSl的主要特征是水的光解和氧气的释放,而PSI则涉及NADP+的还原。光合电子传递链中,光台链上的PC(质体)是一种含铜的电子传递体,而Fc(铁氧还蛋白)是一种含铁的电子传递体,它们在电子传递过程中扮演重要角色。每释放一个氧分子,会有四个电子进入光台电子传递链。
三、光合作用的暗反应
暗反应,又称光合碳循环或Calvin循环,发生在叶绿体基质中.
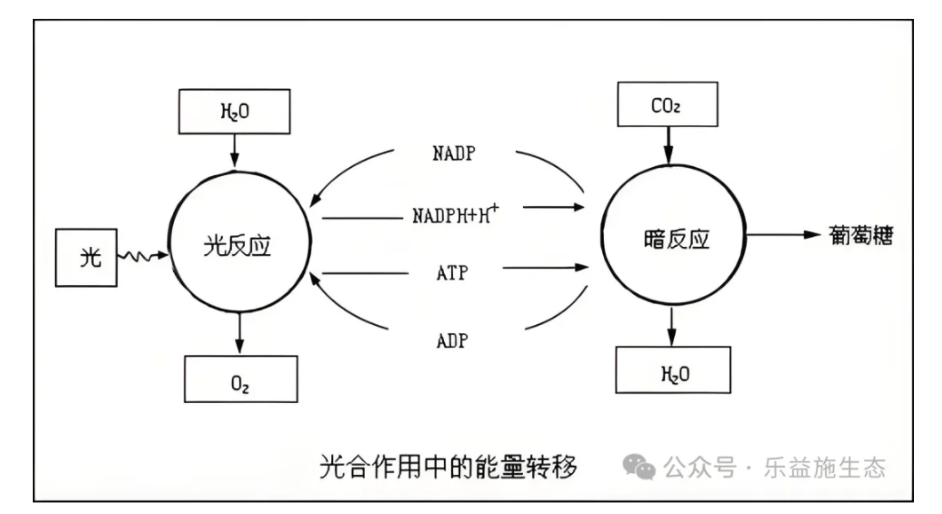
该过程由M,Calvin和Benson提出,他们利用14C同位素标记和双向纸层析技术阐明了C3循环的途径。Rubisco酶在光合作用中扮演着至关重要的角色,它由核基因编码的大亚基和叶绿体基因编码的小亚基组成,活化时需要Mg2+离子的参与。光合碳循环最初形成的糖是三碳糖,而光合作用的主要产物以淀粉和脂肪为主,其中淀粉主要在维管束鞘细胞的叶绿体基质中形成和储存。
四、不同植物的光合作用适应性
光合作用在不同类型的植物中展现出适应性变化。例如,C4植物如玉米和高粱,以及CAM植物如仙人掌和菠萝,发展出了独特的光合机制来提高水分利用效率。
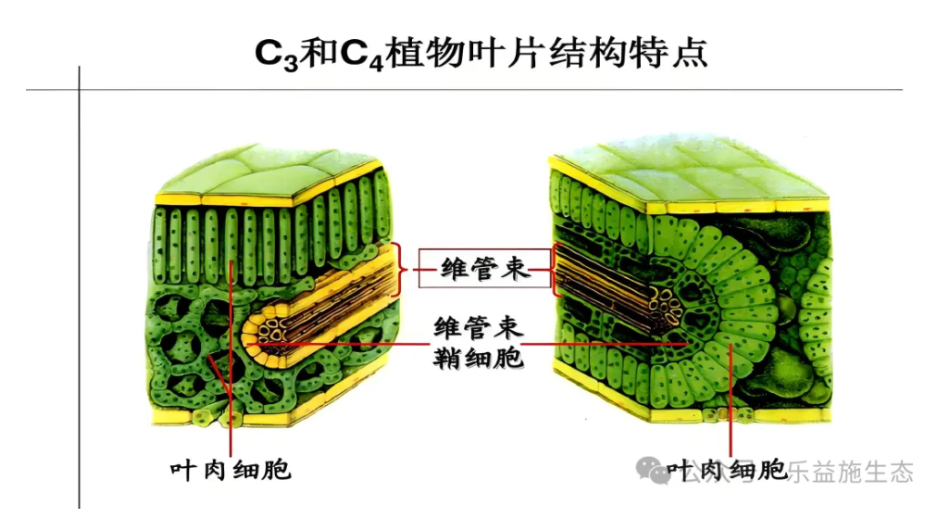
C4植物通过一种称为C4二羧酸途径的过程,先在叶肉细胞中固定C02,然后将其运输到维管束鞘细胞中进行还原,这有助于减少光呼吸损失。CAM植物则在夜间开放气孔吸收C02,并将其周定为苹果酚储存在波泡中,白天再释放C02用于光合作用,从而避免了局温和干燥条件下的水分蒸发。
五、酵母源氨基酸多肽刺激剂促光合作用机理分析
酵母源氨墓酸多肽刺激剂在现代农业中发挥着日益重要的作用,尤其是在提升作物的光合作用效率方面。这类生物刺激剂富含氨基酸、多肽以及其他有益的生物活性物质,它们能够通过多种途径优化作物的光合作用性能,从而促进作物的生长和产量提升。
理论作用机理
营养补充与促进生长:
氨基酸和多肽是构成蛋白质的基本单元,它们可以直接被作物吸收,提供作物生长所需的氮源和其他营养元素,促进叶片的扩展和增厚,从而增加叶面积和叶绿素含量,为光合作用提供更多有效的光合器官。
增强光合器官活力:酵母源氨基酸多肽刺激剂中含有多种氨基酸,特别是谷氨酸和天冬氨酸,这些氨基酸是叶绿体中光合作用相关酶的重要组成部分。通过补充这些氨基酸,可以加速光合酶的合成,提高光合作用速率。
调节光合作用相关基因表达:
研究表明,氨基酸多肽刺激剂能够上调光合作用相关基因的表达,如Rubisco(核酮糖-1,5-二磷酸羧化酶/加氢酶)大亚基和小亚基基因,以及RuBP羧化酶基因,从而提高光合作用效率。提高作物逆境适应性:氨基酸多肽刺激剂中的抗氢化成分可以减轻逆境条件(如干旱、盐碱、低温)对作物的伤害,保护光台器官免受氧化损伤,维持光合作用稳定进行。
文章来源: 乐益施生态
 你可能喜欢的主页
你可能喜欢的主页




 今日话题
今日话题










 最新资讯
最新资讯




 会邦服务号,扫一扫
会邦服务号,扫一扫
 全球展会报名:添加公众号
全球展会报名:添加公众号

提供来自汽车、油气、宠物、印刷、包装、母婴、消费电子、玩具等60多个行业的社区和参与。
注册登记用户来自172个国家和地区。
我们的使命是通过现场体验将世界凝聚在一起。
From Marketes By marketers For marketers。
关注公众号
在线支持
 鄂ICP备2022017323号
鄂ICP备2022017323号
 鄂公网安备 42018502006386
鄂公网安备 42018502006386
